
L. Martínez-Jacobo a C. Córdova-Fletes a, b R. Ortiz-López a, b F. Rivas cC. Saucedo-Carrasco d A. Rojas-Martínez a, b
a 医学部生化学・分子医学科、b 分子生物学ユニット、 ゲノミクスとシーケンシング、 モンテレーのヌエボレオン自治大学健康科学研究開発センター、 c オクシデンテ総合病院、ハリスコ州保健省、グアダラハラ事務局、および d プライベートオフィス、小児科、サンルイスポトシ、メキシコ
キーワード
アレイCGH ・ 候補遺伝子 ・ Delineation del(7q) ・ 裂手裂足領域
要約
本研究では、G‐バンディングおよびCGH‐SNPアレイにより測定した7q21.3q31.1の生まれつきの新しい欠失の女性患者を提示する。 患者は、精神運動遅滞、先天性重度両側緑内障、口蓋裂、および心臓欠損などの特徴を示した。 マイクロアレイアッセイは、欠失した12.5Mb領域を、指やつま先の奇形に関連がある領域の約88kb下流に示した;従って、患者の最終核型は、46,XX.arr 7q21.3q31.1(96,742,140~109,246,085)×1dnであった。 この女児は、先天性緑内障および7q21.3q31.1領域を包含または重複する欠失を示し、このような臨床的特徴に関連する遺伝子座または遺伝子座の存在を確認する、これまでに記載された4番目の患者を表す。 著者らの結果によれば、7q中間欠失に続発する眼欠損への傾向は、TAC1、HBP1、およびチトクロームP450遺伝子の小さなクラスター(サブファミリー3A)の同時欠失によって引き起こされる可能性がある。 この結論は、機能的役割と発現部位によって裏付けられている。また、TAC1は緑内障に関連する変異を有するMYOC遺伝子の機能経路に関連しているためである。 さらに、この女児が7q21q32内の多様な欠失に関連するいくつかの表現型を臨床的に想起させることを考えると、我々の結果および観察は、7q21.3q31.1に重複する欠失/表現型の遺伝子含量の一般的な概観を提供し、CUX1遺伝子およびDLX5から下流の潜在的調節エレメントを含むDLX遺伝子より遠位の座位が、指やつま先の奇形とは無関係であることを確認する。
著作権 2013 S. Karger AG, Basel
はじめに
先天性7q欠失はかなり一般的であるが、不均一である。 これらの欠失は全体として7q(DECIPHERデータベース)を含み、通常、近位(q11→21)、中間(q21→31/32)および末端(q32→qter)に分類される[ギブソン他、1982、ヤング他、1984、チョン他、2008]。
L.M.-J.とC.C.-F.はこの研究に同等に貢献した。7q21q32セグメント内の多様な欠失は、しばしば、指やつま先の奇形および複数の臨床的特徴、例えば、知的障害/発達遅延、耳/聴覚異常、低出生体重、摂食障害、異常な啼泣、小頭症、小顎症、心臓および口蓋欠損、再発性感染、異常な手掌しわ、および眼の異常と関連している[ヤング他、1984、リビエラ他、1991、Scherer他、1994、モントゴメリー他、2000、Bernardini他、2008、チョン他、 欠失領域の見かけ上の重複と共にこの表現型の一貫性は、実際には認識可能な欠失症候群を表している可能性がある[Fagan他 1989]。 本稿では、中間(q21→31/32)欠失に関連する眼の表現型のマッピングを精緻化し、さらなる洞察を提供するために、7q21.3q31.1欠失および先天性重度緑内障(指やつま先の奇形を伴わない)を有する女児について報告する。
患者記述
この患者は1歳の女の子で,血縁関係のない両親から生まれた2番目の子供である。家族歴には異常がなかった。 胎児発育遅延、短大腿骨および胎盤の石灰化が妊娠後期に検出され、彼女は胎児リスクの増加により妊娠35週に帝王切開により出生した。 出生時体重は2,300g(<3パーセンタイル)、身長は43cm(<3パーセンタイル)であり、言及されていたのはアプガースコア8点(10点満点中)のみであった。先天性緑内障は2回別々に流出路再建術(トラベクロトミー、線維柱帯切除術)により診断され、治療された。 患者はいまだに高眼圧のままである。 頭部MRIでは、クモ膜下水腫、髄鞘形成過程の変化、両側視神経浮腫がみられた。 7か月齢時、彼女の体重は4,300g(10~25パーセンタイル)で、身長は57.3cm(<3パーセンタイル)であった。 彼女は大きな眼、青い強膜、大泉門、小泉門の開大、突出した眉部、過剰に折れ込んだ耳輪を伴う低位耳介、外側に挙上した眼瞼裂(上がり目)、明らかな眼角隔離、広い鼻尖、鼻翼の発育不全、鼻翼の低形成、短い鼻柱、両側の片側手掌裂、両側第5指尖部を呈した(図1A)ーーー>鼻底と正中線上唇縁が離れている、口唇が薄い、二重弓形の上くちびる(線)、軟口蓋裂、小顎症、両側の猿線、側方に彎曲している両側第5指尖部がみられた(図1A) また、卵円孔開存、動脈管開存、三尖弁逆流を認めた。満1歳時、頚定していない。 聴覚刺激に対する反応は弱いが、彼女は光刺激に従う。 歯の発達はなく、泣き声も弱く、体重、身長、頭部前後径周囲 はそれぞれ5,500g(3パーセンタイル未満)、67.5cm(<3パーセンタイル、42cm(<3パーセンタイル)であった。 眼科学的検査では、眼球突出、眼圧上昇、青色強膜、角膜混濁を伴う角膜浮腫を特徴とする両側性の原発性先天性緑内障がみられた。 虹彩を含むブドウ膜構造は、明らかに正常であったが、虹彩上に着床して虹彩の血管から血流を受けているかはわからなかった。 現在、患者は頻回の呼吸器感染症罹患から回復している。
材料および方法
染色体分析
患者とその両親の最初の細胞遺伝学的分析は、72時間のリンパ球培養から得たGTGバンド中期染色体で行った。 インフォームド・コンセントの下に患者および両親から採取したさらなる血液サンプルを用いて、分子研究を実施した。
アレイCGH
最初に、患者とその両親からのゲノムDNAをQiagen Gentra® Puregene Blood core Kitを用いて3mlの末梢血から得た。 中密度マイクロアレイ分析は、Agilent SurePrint G3 Hmn CGH+SNP 4x180Kマイクロアレイキット(中央値25kbの間隔を有する約120,000個のCGHプローブおよび60,000個のSNPプローブを含む)を用いて実施した。 簡単に説明すると、患者とその両親、および性別をマッチさせた対照由来のゲノムDNA(約1mg)を、37℃で2時間、AluIおよびRsaI制限酵素(Promega, Madison, Wisc, USA)で消化し、消化された生成物を、SureタグDNA標識キット(Agilent Technologies)を用いてCy3-dU 標識された生成物を精製し、ハイブリダイズさせ、Agilentプロトコルに従って洗浄した。 各スライドをNimblegen MS 200スキャナー(Roche)上でスキャンし、得られた画像を画像変換ソフトウェアで変換し、Feature Extractionソフトウェア(Agilent技術)で画像化した。 結果は、Agilent Cytogenomicsソフトウェアv.2.5によるADM-2収差アルゴリズムを用いて、デフォルト分析法-CGH+SNP v2を用いて分析した。
結果
患者のGバンド核型は46,XX,del(7)(q22q22)dnであり、親核型は正常であった。 対応するマイクロアレイアッセイは、500のマーカーおよび229の遺伝子(miRNAおよび仮想蛋白質を含む)を包含する欠失12.5-Mb領域(ゲノム位置96,742,140-109,246,085)を示し、ACN9はより近位であり、C7orf66はより遠位であった(GRCh37/hg19);従って、7q21.3q31.1に対する部分的モノソミ この欠失は、欠失指形質のオリジナルクリティカル領域[Scherer他、1994]と重なり合っているが、このような臨床的特徴の主要候補であるDLX5およびDLX6遺伝子は含まれていない。 他にゲノムの顕著な変化は観察されなかった。 両親からのマイクロアレイの結果は正常であった。 患者の最終核型は46,XX.arr 7q21.3q31.1(96,742,140~109,246,085)×1dnであった。
考察
本研究では、q21.3q31.1領域を含むまたは重複する生まれつきの新しい欠失を伴った先天性緑内障の世界第4例目の患者を報告した。
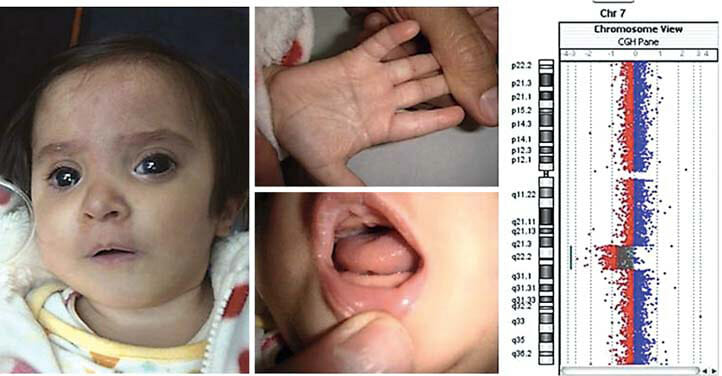

図1
A 顎顔面、手、口 患者さんの特徴。
B ロスプロファイル解析 log 2 ratioで示された(プロットと線)で表示します。ソフトウェアv.2.5で可視化しています。
C 図(ノンスケール 眼球異常がある場合とない場合と 眼球異常がある場合とない場合、および7q21.3q31.1に重なる7q欠失の保因者を示す。7q21.3q31.1に重なる7q欠失の保因者を示しています。ほとんどの これらの染色体異常のほとんど は分子的に精緻化されていないため、我々は これらの染色体異常のほとんどは、分子的に精緻化されていないため、もともと割り当てられていたブレイクポイント もともと割り当てられていたブレイクポイントから近似しています(つまり、私たちは つまり、サブバンドではなく、バンドの近位または遠位の端に サブバンドではなく、バンドの近位または遠位の端にブレイクポイントを配置しています)。)
現在のケース」以下のケースは は、DECIPHERの症例の中で、今回の欠失と主に重なるものである。現在の削除部分と重なっている。縦の 青い線は、眼球異常の潜在的最小領域を示す。眼球異常の潜在的な最小領域を示す。最初の青色の線(左)と点線の 線は、我々の症例と重なる領域を示している。を示している。灰色の線は重なっている一部の領域を 灰色の線で区切られている。* この症例は 眼球異常も外反母趾も報告されていない。外反母趾も報告されていません。7q21q31であった)。) * * 不正確なブレイクポイント (外反母趾なし)。? = 不明な眼球 胎児が剖検されなかったため、眼球への影響は不明。
D UCSCブラウザのスクリーンショット DECIPHERとISCAで報告された全てのインバランスケースが DECIPHERとISCAで報告された7q21.3q31.1領域に重なる 7q21.3q31.1領域に重なる、DECIPHERとISCAで報告されたすべてのインバランス症例を含むスクリーンショット(赤=欠失。青は増幅)。) ISCAの2つの欠失 (IDがnssv578163とnssv582318である。) 7q21.3q31.1領域に完全にまたがっているように見えるが 完全に7q21.3q31.1にまたがっているように見えますが、実際には 7番染色体のモノソミーに対応している。
表1. 7q21q32内で報告された欠失症例からの臨床的特徴であるが、7q21.3q31.1領域と明らかに重複している
| 臨床的特徴 | 症例/参考文献 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15* | 16 | 17 | 今回の事例 | |
| 一般 | ||||||||||||||||||
| 低出生体重児 | – | + | – | – | + | – | – | + | – | + | + | + | + | + | + | – | – | + |
| ID/DD | + | + | + | + | + | + | + | + | + | + | +a | + | + | +? | +b | + | + | + |
| 発育遅延/低身長 | ||||||||||||||||||
| 伸長 | – | – | – | + | + | + | – | + | – | + | – | + | – | + | – | – | – | + |
| 言語遅延 | – | – | – | – | + | + | + | – | – | – | – | – | – | – | – | – | ? | + |
| 性低血圧 | – | + | – | + | + | – | – | – | – | + | – | – | + | – | – | – | – | + |
| 脳波異常/発作 | – | + | – | – | + | – | – | + | – | + | + | – | – | – | – | – | – | – |
| 再発性感染症 | + | + | – | – | + | – | – | – | –? | – | – | – | – | – | – | –? | – | + |
| 頭部/眼窩 | ||||||||||||||||||
| 周囲の広範囲/顕著な額 | + | + | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | + |
| 小頭症 | + | + | – | – | + | – | – | – | – | – | – | + | – | + | – | – | + | – |
| 上向きの傾斜 | – | + | – | – | – | – | – | – | – | – | + | – | – | – | – | – | – | + |
| 表情筋のひだ | – | – | – | + | – | – | – | + | – | – | – | – | – | – | – | – | – | NA |
| 隆起した眉毛 | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | + |
| Hypertelorism/眼角開離 | – | – | – | – | – | – | – | – | – | – | + | – | + | – | + | –? | + | apparently |
| 耳・聴力 | ||||||||||||||||||
| Low set | + | + | – | + | – | – | – | – | – | – | + | – | + | + | – | – | + | + |
| 奇形 | + | + | – | + | + | + | + | – | – | – | + | + | + | – | – | – | – | + |
| 聴覚障害・聴力障害 | – | – | – | – | – | – | – | – | – | – | – | – | + | – | – | –? | – | apparently |
| 鼻 | ||||||||||||||||||
| 扁平/広鼻ブリッジ | – | – | – | – | – | – | – | – | – | – | – | – | – | – | + | – | – | – |
| 球根状の鼻先 | + | – | – | – | + | – | – | + | – | – | + | – | – | – | – | – | – | + |
| 眼球の異常 | + | + | G | – | – | + | – | + | + | + | G | + | + | G | ? | –? | – | G |
| 口 | ||||||||||||||||||
| 大 | + | + | – | + | + | – | + | + | – | + | + | – | + | – | – | – | – | – |
| 薄い上唇 | – | – | – | + | – | – | – | + | – | ? | ? | – | ? | – | – | – | – | + |
| 長い人中 | + | – | – | – | – | – | – | – | – | + | – | – | – | – | – | – | – | apparently |
| 口蓋裂・高口蓋・狭口蓋 | – | – | + | + | – | – | – | – | – | – | – | + | – | + | – | – | – | + |
| 小顎症 | + | + | – | – | – | – | – | – | – | + | – | + | + | – | – | – | – | + |
| 頸部の栄養障害 | – | + | – | – | + | + | + | + | – | + | + | – | + | – | – | – | – | mildc |
| 首 | ||||||||||||||||||
| 短い | – | – | – | + | – | + | – | – | – | – | + | – | – | – | – | –d | – | – |
| 四肢・手 | ||||||||||||||||||
| 第5指の無指症 | – | + | – | – | – | – | mild | + | – | – | – | – | – | – | – | – | + | + |
| 分裂手指/分裂足 | – | – | – | – | – | – | – | – | – | – | – | + | + | + | – | – | – | – |
| 掌底のしわの異常 | – | + | – | – | + | – | – | + | – | – | + | – | – | – | + | – | – | + |
| 心血管 | ||||||||||||||||||
| ASD/VSD | – | + | + | – | – | – | – | – | – | – | + | – | + | + | – | – | – | – |
| PDA | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | + |
| 肺動脈狭窄 | – | – | – | – | – | – | – | – | – | + | – | – | – | – | – | + | – | – |
| その他 | – | – | – | – | – | – | – | – | – | + | + | – | – | – | + | – | – | +e |
| 消化管 | ||||||||||||||||||
| ヘルニア | – | – | – | – | – | – | – | + | – | – | – | + | + | – | – | – | – | – |
| 泌尿器系 | ||||||||||||||||||
| 性器異常 | – | – | + | – | – | + | – | + | – | – | – | + | – | + | – | + | – | – |
1 Ayraud et al. [1976]; 2 Higginson et al. [1976]; 3 デニス他 [1977]; 4 Hull et al. [1979]; 5 Klep-de Pater et al. [1979]; 6 Serup [1980]; 7 Abuelo and Padre-Mendoza [1982]; 8 ヤング他 [1984]; 9 Chitayat et al. [1988]; 10 Fagan et al. [1989]; 11 Franceschini et al. [1978]; 12 Tajara et al. [1989]; 13 Morey and Higgins [1990]; 14 Montgomery et al. [2000]; 15 Cheong et al. [2008]; 16 ID 253694; 17 ID 255298.
ID/DD = 知的障害/発達遅延。 NA = 利用できない。 G = 緑内障。 ? =不明/不正確。
22週目に出産した胎児で、これ以上の評価はできない。 a 私たちが提案したもの(早期死亡)。 b 著者が提案したもの(早期死亡)。 c 最初は吸い付きが悪かった。 d この患者は手に前軸多指症を呈していた。
このレビューの後、著者らの症例と非常に類似した欠失が24週齢の胎児で報告され、口唇裂と口蓋裂、高色素症、広範な鼻ブリッジ、小顎症、低位耳、小陰茎および停留精巣を呈した[チェン他 2013]。 この表では、症例の30%を超える特徴として、ID/DD(~18/18)、眼奇形(12/18)、低出生体重(10/18)、耳奇形(10/18)、心臓奇形(9/18)、大口(9/18)、摂食障害(9/18)、低設定耳(8/18)、発育遅滞/低身長(8/18)、筋緊張低下(6/18)、小頭症(6/18)、小顎 「弱い泣き声」または「異常な泣き声」という特徴は、本表には含まれていないが、本表を含めて約5例の患者に認められた。
(図1C、表1)を示す。 著者らの結果は、この領域でマッピングされた少なくとも3つの眼関連遺伝子座(TAC1、CYP3A43およびHBP1)の共欠失が緑内障および他の眼の異常を説明できることを示唆する。
臨床的および遺伝的に不均一な状態である緑内障は、網膜神経節細胞の喪失および視神経の萎縮を特徴とする[イゾッティ他 2011; Mookherjee et al., 2012]。 眼圧上昇の主な臨床基準は、通常、房水に対する小柱網の抵抗性に起因する[ケネディ他 2012]。 小柱網は角膜底部の周囲に位置し、房水流出の調節に重要な役割を果たしている[イゾッティ他 2011]。 先天性緑内障患者では、小柱網の変化が頻繁に認められている。 先天性緑内障、濁った角膜、原発開放隅角緑内障、および若年性開放隅角緑内障は緑内障のサブタイプの一部であり、GLC1AまたはMYOC(1q24.3)、CYP1B1(2p22.2)、およびCAV1/CAV2(7q31.2)[Stoilov et al.、1997;Alward et al.、1998;Thorleifsson et al.、2010;ケネディ他、2012;
CYP1B1遺伝子は一次コンジェントと連鎖している
角膜の透明性および房水分泌を調節するチトクロームP450依存性代謝産物を伴う緑内障(OMIM 231300)。 さらに、CYP1B1はダウンレギュレートされ、変異MYOC遺伝子を持つヒト小柱網細胞においてCYP26B1がアップレギュレートされた[ケネディ他、2012]。 これらの事実は、チトクロームP450sファミリーの他の遺伝子も同様に関連している可能性があることを示唆している。 実際には、7q22.1(CYP3A4、CYP3A5、CYP3A7、CYP3A43)にあるチトクロームP450遺伝子の小さなクラスター
実際、我々の患者では、緑内障、または混濁角膜、巨大角膜、または異常瞳孔などの他の眼の異常に関連する7qの欠失のほとんどがq22に関与し、おそらくCYP遺伝子が関与していた[Young他、1984;Montgomery他、2000]。 注目すべきことに、CYP3A43はヒト角膜上皮組織において差次的に発現されることが認められた[ターナー他 2007]。 一般に、サブファミリー3Aのメンバーは、ヒト虹彩、毛様体および角膜において発現されることが認められた[チャン他、2008;ヴォロチネン他、2011]。 緑内障のもう1つの候補遺伝子はTAC1であると思われ、これも我々の患者では欠失しており、最近緑内障損傷の生理学的バイオマーカーとして可能性が示唆されている。 TAC1遺伝子は、MYOC遺伝子の機能経路に関連していた。MYOC遺伝子は、小柱網におけるストレス条件によって誘導される糖蛋白質である。 実際、TAC1発現もMYOC突然変異細胞において強く変化した[ケネディ他 2012]。 TAC1遺伝子(7q21.3)はprecur-をコードする
神経伝達物質として作用するホルモンのソール(UCSCゲノムブラウザー)は、ヒト線維柱帯の網目構造において機械感受性遺伝子として同定されている[ケネディ他 2012]。
この患者でも欠失している別の遺伝子、すなわちHBP1は、WNT経路に関与し、網膜、角膜および毛様体(EMBL-EBIおよびGeneCardsデータベース)に発現する転写抑制因子である。 WNT経路は眼圧調節に関連している[ケネディ他2012]。 同時に、HBP1(高移動度グループボックス転写因子)遺伝子は、網膜損傷および炎症ストレスのシグナル伝達のための内因性分子であることが知られているHMGB1(高移動度グループボックス1タンパク質)と同様である[Lee他、2012]。
7q21q32欠失症の表現型は多岐にわたるが、いくつかの再発性の臨床的特徴が観察されている(表1)。表1で得られた情報によると、少なくとも14の特徴が30%以上の頻度で患者に見られた(表1)。例えば、今回の子どもは、知的障害や発達遅延、成長遅延、頭蓋顔面・心臓・眼球の欠損など、これらの特徴のうち少なくとも11項目を示していました。さらに、ブレイクポイント解析により、今回の欠失の近位ブレイクポイントがDLX5遺伝子のわずか88kb下流にあることが明らかになった。DLX5およびDLX6遺伝子のハプロ不全は外反母趾の原因とされていることから[Scherer他, 1994; van Silfhout他, 2009]、このような四肢欠損を決定する上で、DLX5およびACN9の下流(RP11-800O14とD7S618の間)にあるCUX1遺伝子(Bernardini他[2008]により示唆されている)および潜在的な制御要素(Tzschach他[2007]により示唆されている)が除外されることが間接的に確認された。
以上の結果から、7q21.3q31.1が脳、心臓、成長、眼の生理・発達に重要な遺伝子の多い領域であることが確認され(表2)、7q21.3q31.1に重なる欠失・表現型の遺伝子内容の概要が明らかになり、DLX遺伝子の遠位にある遺伝子座は外反母趾とは無関係であることがさらに確認された。
表2. 7q21.3q31.1の欠失遺伝子は、他の類似の欠失症例でみられるいくつかの頻繁な臨床的特徴に関連する可能性がある
| 関連 | 遺伝子/タンパク質 | 参考文献 |
|---|---|---|
| 脳/ID/DD | ACHE、ATXNL7、BHLHA15、COG5、GPC2、MLL5*、NPTX2、NRCAM、PNPLA8、RELN、SRPK2、SYPL1、TAC1、 THAP5、TMEM130、VGF、NYAP1a | UCSCゲノムブラウザ、Al-Hassnan他[2011] Rymen他[2012]、ID263273を解読する。 ビンセント他[2008]、Uliana他[2010] |
| 頭蓋顔面の変化 | MLL5*、PLOD3**、RELN | Al-Hassnan他[2011]、Salo他[2008] |
| 難聴 | GJC3、LHFPL3、MLL5*、PLOD3**、SLC26A4、SLC26A5 | UCSCゲノムブラウザ、ID263273、Salo他[2008] アルバート他[2006] |
| 眼の異常 | CLDN15、CYP3A4、CYP3A43、CYP3A5、CYP3A7、HBP1、NRCAM、TAC1、TMEM130 | EMBL-EBI、ヴォロチネン他[2011]、リー他[2012]、デミヤネンコ他[2011]、ケネディ他[2012] |
| 心臓/心臓の欠陥 | MOSPD3、NPTX2、PNPLA8、SRPK2、THAP5 | UCSCゲノムブラウザ、ポール他[2004] |
| 消化管の変化/発育遅延・低身長 | CLDN15、MUC3A、MUCB、MUC17、MOGAT3 | UCSCゲノムブラウザ、ワダ他[2013] |
これらの関連性は、遺伝子機能および発現位置、他の症例との遺伝子/表現型の重複、または以前の関連に従って実施された。 いくつかの遺伝子は、複数の臨床的特徴に関連している可能性があることに留意されたい。 実際、いくつかの単遺伝子性病変が様々な臨床的特徴(例)に関与しているように思われた。 MLL5*またはPLOD3**。
a ISCA微小欠失例(ID nssv578204)で削除。
謝辞
患者の両親の継続的な協力とDr. Jose A. Paczka(眼科医)の報告に感謝申し上げます。 この研究はPROMEP(No)によって部分的に支援された。 103.5/11/4330, PAICYT (SA609-10) to C. Cordova-Fletes, PAICYT (No.) SA324-10、FOMIX(Convocatoria M0014-2007-2010、Reg) 068251) CIDICS L. Martinez-Jacoboは、CONACYT奨学金によって支援されている。
参考文献
Abuelo DN, Padre-Mendoza T: 7qの間質欠失によるCat様泣きおよび精神遅滞(7q22は7q32につながる)。 J Med Genet 19: 473-476 (1982)
Albert S, Blons H, Jonard L, Feldmann D, Chauvin P, et al: SLC26A4遺伝子は白人集団における前庭水管肥大を伴う非症候性聴覚障害にしばしば関与している。 Eur J Human Genet 14:773-779 (2006)
Al‐Hassnan ZN、Al‐Bakheet A、Abu‐Dheim N、Al‐Younes B、Colak D、Kaya N:アレイ比較ゲノムハイブリダイゼーションにより検出された7q22.1~7q22.3の新規間質微小欠失。 Am J Med Genet 155:3128-3131 (2011)
Alward W, Fingert J, Coote M, Johnson A, Lerner S, et al: 第1染色体開放隅角緑内障遺伝子(GLC1A)の突然変異と関連する N Engl J Med 338:1022-1027 (1998)
Ayraud N, Rovinski J, Lambert JC, Galiana: 第7染色体長腕の間質性欠失。 Ann Genet 19:265-268 (1976)
Bernardini L, Palka C, Ceccarini C, Capalbo A, Bottilo I, et al: 染色体7q21.13-q22.1の複雑な再編成は、直指難聴遺伝子座を確認し、新しい候補遺伝子を示唆する。 Am J Med Genet A 146A:238-244 (2008)
Chen CP, Chang SJ, Chern SR, Wu PS, Chen YT, et al: 7qのde novo interstitial deletement(7q22.1→q31.1)の出生前診断および 遺伝子521:311-315(2013)
Cheong MLJ, Tsai MS, Cortes R, Harrison MR: 第1トリメスターダウン症候群スクリーニングで検出された染色体7qの中間的間質欠失。 胎児診断24:340-344(2008)
Chitayat D、McGillivray BC、Wood S、Kalousek DK、Langlois S、Applegarth DA:間質性7q欠失[46,XX,del(7)(pter→q21.1::q22→qter)]およびβグルクロニダーゼおよび嚢胞性線維症の遺伝子の位置 Am J Med Genet 31:655-661 (1988)
Demyanenko G, Riday T, Tran TS, Dalal J, Darnell EP, et al: NrCAM欠失は視床皮質軸索を視覚皮質にトポグラフィー的な誤配向させ、視力を破壊する。 J Neurosci 31:1545-1558(2011)
Dennis NR, Neu RL, Bannerman RM: 多発性奇形の乳児における部分的7qモノソミー。 Am J Hum Genet 29:37A (1977)
Fagan K, Gill A, Henry R, Wilkinson I, Carey B: 7qの間質性欠失およびβ-グルクロニダーゼ遺伝子の排除マッピングの要約。 J Med Genet 26:619-625 (1989)
Franceschini P, Silengo MC, Davi GF, Santoro MA, Prandi G, Fabris C: 染色体7 46,XX,del(7)の長腕の間質性欠失(pterはq2200:q32 Hum Genet 44:345-348 (1978)
Gibson J, Ellis PM, Forsyth JS: 第7染色体の間質性欠失: 症例報告および文献レビュー。 Clin Genet 22:256-265 (1982)
Higginson G, Weaver DD, Magenis RE, Prescott GH, Haag C, Hepburn DJ: 多発性奇形の乳児における染色体7番(7q-)の長腕の間質性欠失。 Clin Genet 10:307-312 (1976)
Hull DR, Kessler KK, Juberg RC: 7qの間質性欠失により、発育不全および特異的な泣きを引き起こす: 以前に報告された7q1および7q2の欠失との比較。 Am J Hum Genet 31:97A (1979)
Izzotti A, Longobardi M, Cartiglia C, Sacca SC: 小柱網におけるミトコンドリア損傷は、原発性開放隅角緑内障および偽剥脱性緑内障においてのみ起こる。 PLoS 1 6e14567 (2011)
Kennedy KD, Anithachristy SA, Buie LK, Borras T: Cystatin A: 変異型ミオシリン原因緑内障の一般的な関連の可能性がある。 PloS 1 7:e36301 (2012)
Klep-de Pater JM, Bijlsma JB, Bleeker-Wagemakers EM, de France HF, de Vries-Ekkers CMAM: 第7染色体長腕の異なる欠失を有する2症例 J Med Genet 16:151-154 (1979)
Lee J, Hsiao C, Yang I, Chou MH, Wu CL, et al: High-mobility group box 1蛋白質は、ラット網膜神経節細胞株RGC-5における糖化最終産物誘導性血管内皮増殖因子A モールビジョン18:838-850(2012)
Montgomery TL、Wyllie J、Oley C:7q21.2-q31.2の間質性欠失に関連する乳び指症および緑内障。 Clin Dysmorphol 9:235-239 (2000)
Mookherjee S, Acharya M, Banerjee D, Bhattacharjeee A, Ray K: MYOCアップレギュレーションにおけるCYP1B1の関与の分子的根拠および緑内障の病因におけるその潜在的意義 PLoS 7:e45077 (2012)。
Morey MA, Higgins RR: 7qの間質性欠失を伴うエクトロ-アメリア症候群。 Am J Med Genet 35:95-99 (1990)
Pall GS, Wallis J, Axton R, Brownstein DG, Gautier P, et al: 右室発達に役割を果たす新しい膜貫通MSP含有蛋白質。 ゲノム84:1051-1059(2004)
Rivera H, Sanchez-Corona J, Burgos-Fuentes VR, Melendez-Ruiz MJ: 7q22の欠失と欠失指数。 Genet Couns 2:27-31 (1991)
Rymen D、Keldermans L、Race V、Rregal L、Deconinck N, et al: COG5-CDG: 臨床スペクトルの拡大。 孤児院J Reare Dis 7:94 (2012)
Salo AM, Cox H, Farndon P, Moss C, Grindulis H, et al: リシルヒドロキシラーゼ3遺伝子の突然変異によって引き起こされる結合組織障害。 Am J Hum Genet 83:495-503 (2008)
Scherer S, Poorkaj P, Allen T, Kim J, Geshuri D, et al: 染色体7上の常染色体顕性スプリントハンド/スプリットフット遺伝子座のファインマッピング、バンドq21.3- 1. Am J Hum Genet 55:12-20 (1994)
Serup L:第7染色体長腕の間質性欠失。 Hum Genet 54:19-23 (1980)
Stoilov I、Akarsu N、Sarfarazi M:染色体2p21上のGLC3A lo-cusに関連する家系における原発性先天性緑内障(ブドウ球菌)の主要原因としてのチトクロームP4501B1(CYP1B1)における3つの異なる切断突然変異の同定。 Hum Mol Gen 6:641–647 (1997)
Tajara EH, Varella-Garcia M, Gusson AC: 第7染色体の間質性長腕欠失および直指症。 Am J Med Genet 32:192-194(1989)
Thorleifsson G, Walters GB, Hewitt AW, Masson G, Helgason A, et al: CAV1およびCAV2に近い一般的な変異型は、原発開放隅角緑内障 Nat Genet 42:906-909 (2010)
Turner H、Budak M、Akinci M、Wolosin JM:オリゴヌクレオチドマイクロアレイを用いたヒト結膜および角膜上皮遺伝子発現の比較分析。 Ophthalmol Vis Sci 48:2050-2061 (2007)に投資する。
Tzschach A, Menzel C, Erdogan F, Schubert M, Hoeltzenbein M, et al: tiling path array CGHによる16 Mbの間質性染色体7q21欠失の特徴 Am J Med Genet A 143:333-337 (2007)
Uliana V, Grosso S, Cioni M, Ariani F, Papa FT, et al: 染色体7のバンドq22.2-q22.3における3.2Mbの微小欠失は、過成長および遅延 Eur J Med Genet 53:168-170 (2010)
van Silfhout AT, van den Akker PC, Dijkhuizenet T, Verheij J, Olderode-Berends M, et al: 染色体7q異常(SHFM1)による手足のスプリット奇形: 原因機序としての機能的半 Eur J Hum Genet 17:1432-1438 (2009)
自閉症および精神遅滞に関連するビンセントJB、Choufani S、Horike S、Stachowiak B、Li M他:A転座t(6;7)(p11-p12;q22):切断点における候補遺伝子の局在および同定。 精神科Genet 18:101-109 (2008)
Volotinen M, Hakcola J, Pelkonen O, Vapaatalo H, Mmaenpa A J: 眼科用チモロールの代謝: 古い薬の新しい側面 基本クリン薬理学ツール108:297-303 (2011)
和田M, 田村A, 高橋N, 津北S: マウスからのクローディン2および15の喪失は、腸の傍細胞性Na(+)流および栄養輸送の欠損を引き起こし、栄養失調による死亡をもたらす。 消化器内科:S0016-5085(2012)
Young RS, Weaver DD, Kukolich MK, Heerema NA, Palmer CG, et al: 第7染色体長腕の末端欠失および間質欠失: 新たに5例を対象とした総 Am J Med Genet 17:437-450 (1984)。
Zhang T, Xiang C, Gale D, Carreiro S, Wu E, Zhang EY: 薬物輸送体およびヒト眼関門におけるチトクロームP450 mRNA発現: 眼内薬物動態への影響 ドラッグ・メタブ・ディスポ36:1300-1307(2008).
KARGER
e-Mail karger@karger.com ww.karger.com/msy著作権 2013 S. Karger AG, Basel 1661-8769/13/0046-0285$38.00/0
オーガスト・ロジャス・マルティネス、MD/DSc
分子生物学連合、ゲノミカ、科学研究センター、サウード市民団体、共同研究機関
モンテレイ64460 (メキシコ) ヌエボ・レオン大学電子メール・アロジャストムツ@gmail.com
日本皮膚科学会 皮膚科専門医/日本医師会 産業医/東京衛生検査所 指導監督医
この記事は、 ヒロクリニックNIPTの編集・監修体制 にもとづき、資格を持つ医師が内容を確認しています。


