
この記事のまとめ
猫なき症候群は、5番染色体の短腕(5p)が一部欠けることで起こる先天的な病気で、乳児期の高い泣き声・発達の遅れ・特徴的な顔立ちなどがみられます。名前は、赤ちゃんの泣き声が子猫の鳴き声に似ていることに由来します。多くは新たに生じた変化で、親から受け継ぐケースは限られます。近年の研究では、欠けた場所によって出やすい症状が異なることが分かってきました。5pの欠失のような構造の変化は、一般的なNIPT(新型出生前診断)の主な対象ではなく、確定診断には羊水検査や絨毛検査と染色体の詳しい分析が必要です。気になることがあれば、遺伝カウンセリングで専門家に相談してください。
この記事でわかること
- 猫なき症候群とは何か、名前の由来になった高い泣き声のこと
- 原因となる5番染色体短腕(5p)の欠失と、その多くが新たに生じる理由
- 泣き声・発達の遅れ・顔立ちなど主な症状と、欠けた場所で症状が変わる研究の知見
- 診断の流れとNIPT・羊水検査の位置づけ、療育や家族の支えになる相談先
猫なき症候群とは|5番染色体短腕(5p)が欠けて起こる病気
猫なき症候群は、5番染色体の短腕(5p)の一部が欠けることで発達に影響が出る、生まれつきの染色体の病気です。5pモノソミー、または5p−症候群とも呼ばれます。生まれてくる赤ちゃんのおよそ1万5千人から5万人に1人ほどとされ、決してありふれた病気ではありません。まずは、どんな病気なのかを整理します。
この病気の名前は、乳児期の泣き声に由来します。生まれてすぐの赤ちゃんの泣き声が、子猫の甲高い鳴き声に似ていることから、こう呼ばれるようになりました。泣き声のほかにも、発達の遅れや特徴的な顔立ちなど、いくつかのサインが重なって現れます。
症状の程度には、大きな個人差があります。同じ病名でも、欠けた部分の大きさや場所によって、現れ方はさまざま。療育や医療の支えを受けながら、その子のペースで成長していくお子さんも多くいます。過度に不安を抱えず、まずは正しい知識を持つことが第一歩です。
名前の由来|乳児期の高い泣き声というサイン
猫なき症候群の最初のサインは、生後間もない時期に聞かれる高くて細い泣き声で、これが病名の由来になっています。喉頭(のど)の発達に関わる特徴が背景にあると考えられています。この泣き声は成長とともに目立たなくなることが多く、乳児期に気づかれやすいサインです。
後で紹介する研究では、この特徴的な泣き声が、5p15.31という遠い側の領域の欠失と関係することが示されました。泣き声というひとつのサインにも、染色体上の位置が対応している。そう考えると、症状の背景を少し具体的にイメージできます。
ただし、泣き声だけで診断が決まるわけではありません。ほかの特徴とあわせて、医師が総合的に判断します。乳児期に気になる泣き声が続くときは、自己判断で様子を見ず、かかりつけの小児科へ相談してください。
原因|5番染色体短腕の欠失と、多くは新たに生じる変化
猫なき症候群の原因は、5番染色体の短腕(5p)の一部が欠けること(欠失)で、その多くは新たに生じた変化(新生突然変異)です。親から受け継いだものではなく、精子や卵子ができる過程、あるいは受精直後に偶然起こることがほとんど。つまり、親の育て方や妊娠中の過ごし方が原因ではありません。
欠け方には、いくつかのパターンがあります。染色体の先端が欠ける末端欠失が多く、途中の一部だけが欠ける間質性欠失もみられます。あとで紹介する患者94人の研究でも、多くが5pの単純な欠失を持っていました。
一方で、ごく一部は親から受け継ぐケースもあります。親が均衡型転座という、染色体の一部が入れ替わっても量に過不足がないタイプの持ち主(保因者)だと、子どもに欠失が伝わることがあるのです。この場合、次の妊娠での可能性を知るうえで、遺伝カウンセリングが助けになります。染色体の微小な欠失全般については微小欠失症候群のリスクを解説した記事もあわせてご覧ください。
主な症状|泣き声・発達の遅れ・顔立ち・成長
猫なき症候群の主な症状は、乳児期の高い泣き声、発達や言葉の遅れ、知的発達の遅れ、特徴的な顔立ち、そして低出生体重や成長のゆっくりさです。すべてが同じ強さで出るわけではなく、お子さんによって現れ方は異なります。ここでは代表的なサインを整理します。
顔立ちの特徴としては、丸顔、両目の間隔が広め、目尻がやや下がる、あごが小さめといった点が挙げられます。これらは乳幼児期に目立ち、成長とともに変化していくことが多いとされています。筋緊張が弱め(筋緊張低下)で、体がやわらかく感じられる場合もあります。
下の表に、主な症状と気づかれやすい時期をまとめました。あくまで一般的な傾向であり、当てはまらないお子さんもいます。診断や見通しは、必ず主治医の説明にもとづいて理解してください。
| 主な症状 | 気づかれやすい時期 | 特徴 |
|---|---|---|
| 高い泣き声 | 新生児〜乳児期 | 子猫のような甲高い泣き声。成長とともに目立たなくなることが多い |
| 発達・言葉の遅れ | 乳幼児期以降 | 運動やことばの発達がゆっくり。療育でペースに合わせて支える |
| 知的発達の遅れ | 幼児期以降 | 程度に個人差が大きい。欠失の大きさや場所と関係する |
| 特徴的な顔立ち | 乳幼児期 | 丸顔・目の間隔が広め・小さめのあごなど。加齢で変化 |
| 低出生体重・成長 | 出生時〜乳幼児期 | 生まれたときの体重が小さめ、成長がゆっくりな場合がある |
発達や知的面の遅れは、療育や医療的な支えを受けながら、その子のペースで伸びていきます。妊娠中に知的発達の遅れが心配な方は、知的障害のリスクについて解説した記事も参考になります。焦らず、専門職と一緒に歩むことが支えになります。
欠失の場所と症状の関係|患者94人の研究からわかったこと
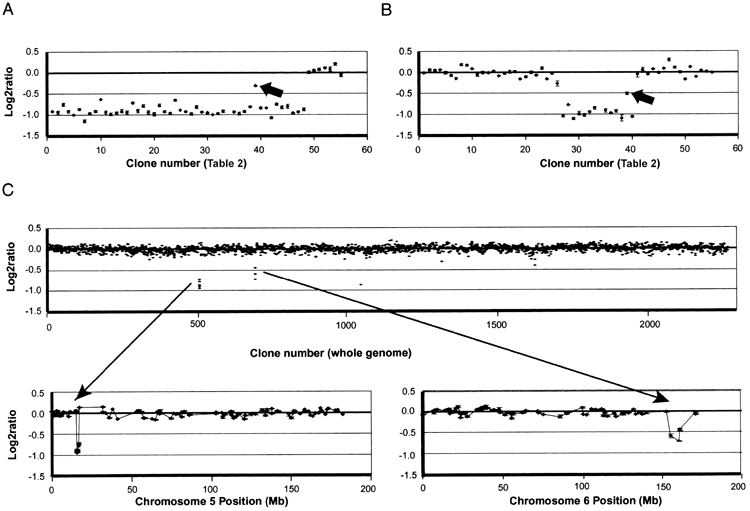
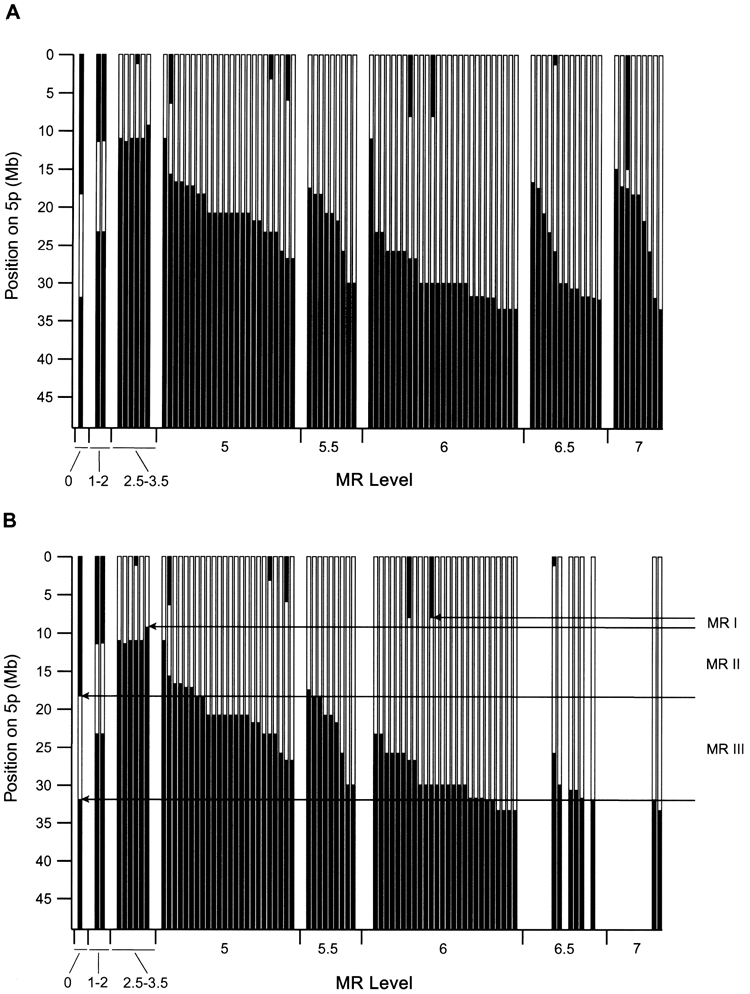
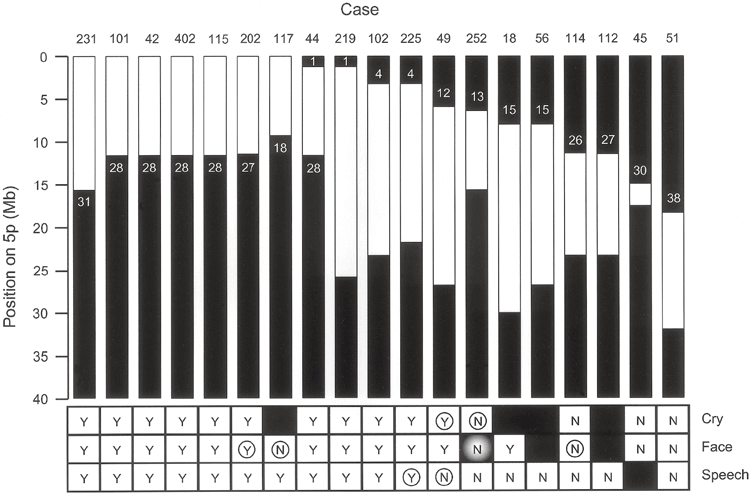
近年の研究では、5pのどこが欠けるかによって出やすい症状が異なり、知的発達の程度は欠失の大きさと位置におおむね依存することが分かってきました。これは、染色体の詳しい解析(アレイ比較ゲノムハイブリダイゼーション)で患者94人を調べた研究の知見です。症状の背景を理解する手がかりになります。
この研究では、特徴ごとに関係する領域が絞り込まれました。私自身、こうしたマッピングの図を初めて見たとき、ひとつの病気の中にこれほど細かな対応関係があることに驚いた記憶があります。症状が「場所」と結びつくと、見通しの立て方も変わってきます。
研究で示された対応を、かみくだいて紹介します。特徴的な高い泣き声は5p15.31付近、言葉の遅れは先端に近い5p15.32〜15.33付近、顔立ちの特徴は5p15.2〜15.31付近の欠失と関連していました。同じ病気でも、欠ける場所で前に出るサインが違ってくるわけです。
知的発達の遅れについては、欠失が大きく、複数の領域にまたがるほど程度が強くなる傾向がみられました。さらに注目したいのが、遅れが特に重かった例です。その約3分の2で、5pの欠失に加えて別の染色体の変化を併せ持っていたことが分かりました。重い遅れの背景には、5p以外の要因が関わることもあるのです。
以下は、この研究で示された遺伝子型と表現型の対応を表した図です。専門的な内容を含みますが、症状が染色体上の位置と丁寧に対応づけられていることが読み取れます。参考資料として掲載します。
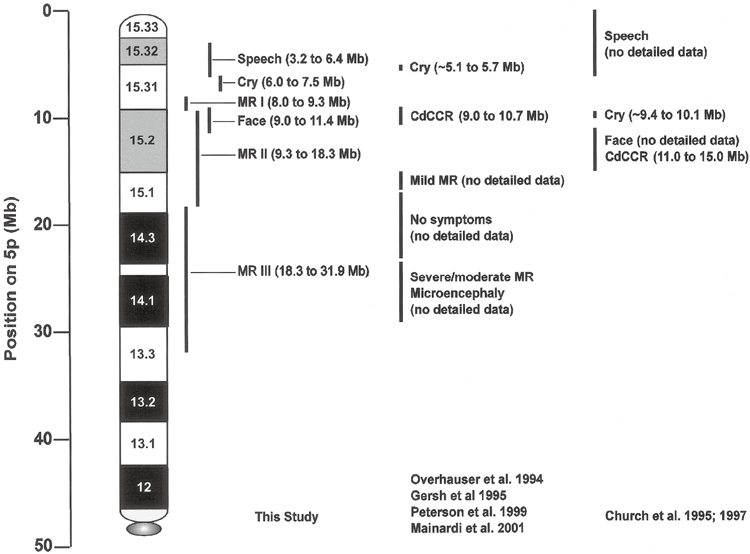
ここで大切なのは、これらが症状の傾向を示すものであって、一人ひとりの見通しをそのまま決めるものではない、という点です。実際の発達は、療育や環境、家族の支えなど多くの要素で変わります。数値や位置に一喜一憂しすぎず、目の前のお子さんの歩みを見つめてください。
診断|染色体検査・マイクロアレイと出生前診断での位置づけ
猫なき症候群の確定診断は、染色体を調べる検査で行い、細かな欠失には染色体マイクロアレイという詳しい解析が用いられます。出生後は、赤ちゃんの症状をきっかけに染色体検査へ進むことが一般的です。まずは、どんな検査で分かるのかを整理します。
ここで押さえておきたいのが、NIPT(新型出生前診断)との関係です。NIPTは、母体の血液中を流れる赤ちゃん由来のDNA(細胞外を流れるDNA)を調べる、非確定的検査。主に13・18・21トリソミーなどの染色体の数の変化を対象にしています。
5pの欠失のような構造の変化は、一般的なNIPTの主な対象ではありません。微小な欠失を一部の施設がオプションで調べる場合もありますが、対象は限られ、検出率にも幅があります。NIPTで何が分かるかはNIPTでわかることを解説した記事でも詳しく紹介しています。あくまでスクリーニングであり、結果が陽性でも確定ではありません。
確定のためには、羊水検査や絨毛検査で採取した細胞を、染色体マイクロアレイなどで詳しく調べます。これらは診断を確定できる一方、わずかながら流産のリスクがあります。目安として、羊水検査でおよそ0.3%前後、絨毛検査でおよそ1%前後と報告されています。検査を受けるかどうかは、メリットと負担を理解したうえでの自己決定です。羊水検査の詳細は羊水検査について解説した記事をご覧ください。
検査との向き合い方|段階的に考え、限界も知っておく
出生前検査は、何をどこまで知りたいかを夫婦で話し合い、段階的に考えることが、納得のいく選択につながります。どの検査も、すべての病気や障害を見つけられるわけではありません。分かる範囲と限界を知っておくと、結果をどう受け止めるかの心の準備ができます。
私が相談の場で感じるのは、「結果が出る前に、夫婦で方針を決めておくかどうか」で迷う方が多いことです。答えはひとつではありません。Xでも@chiisunmonさんが、受けるべきか迷ったけれど結果が陰性で安心材料になったこと、陽性だったときのことはあえて夫婦で決めずに受けた、その時に考えようと思ったとつづっていました。決めておく人もいれば、そうでない人もいる。どちらも、その家庭なりの向き合い方です。
検査を段階的に組み立てる考え方も、負担を抑える一つの方法です。まず初期のスクリーニングを受け、必要に応じて次の検査へ進む。こうした順序を決めておくと、判断がぶれにくくなります。@Hzkn2oCsさんも、まず初期スクリーニングを受けて陽性ならNIPTへと段階を決めていたこと、すべての障害の可能性を排除できるわけではないけれど安心が買えたと思うと話していました。検査で分かる範囲には限界がある、という前提を共有しておくことが大切です。
結果をどう受け止めるか迷ったときは、一人で抱え込まないでください。遺伝カウンセリングでは、専門家が中立の立場で情報を整理し、あなたの気持ちに寄り添います。実際の陽性事例や向き合い方の一例はNIPT検査の体験を紹介した記事も参考になります。
療育と家族の支援・予後
猫なき症候群は、早くから療育や医療的な管理を組み合わせることで、その子のペースに合わせた成長を支えられます。根本的に染色体を元に戻す治療はありませんが、発達を後押しする関わりや、体調面のフォローは数多くあります。ここでは支えになる考え方を紹介します。
乳幼児期には、理学療法や作業療法、言語の支援などが役立ちます。心臓や消化器などに合併症がある場合は、それぞれの専門科が連携してケアします。摂食や睡眠の困りごとも、専門職と相談しながら一つずつ整えていく。地道な積み重ねが、暮らしやすさにつながります。
家族にとって心強いのが、公的な相談窓口や支援情報です。難病や発達に関する情報は難病情報センターで調べられます。発達障害に関する支援は国立障害者リハビリテーションセンター 発達障害情報・支援センターが参考になります。出生前検査全般の考え方は出生前検査認証制度等運営委員会でも確認できます。
予後には幅があります。医療的なケアを続けながら、家庭や地域で穏やかに暮らしているお子さんも多くいます。同じ診断名でも歩みはさまざま。だからこそ、比較や不安に振り回されず、その子の「できた」を一緒に喜ぶ姿勢が支えになります。困ったときは、遠慮なく専門職を頼ってください。
よくある質問
Q. 猫なき症候群は遺伝しますか?
多くは新たに生じた変化(新生突然変異)で、親から受け継いだものではありません。精子や卵子ができる過程などで偶然起こることがほとんどです。ただし、ごく一部は親が均衡型転座の保因者で、そこから欠失が伝わるケースがあります。次の妊娠での可能性が気になる場合は、遺伝カウンセリングで専門家に相談してください。
Q. NIPTで猫なき症候群は分かりますか?
一般的なNIPTは、主に13・18・21トリソミーなど染色体の数の変化を対象とする非確定的検査で、5pの欠失のような構造の変化は主な対象ではありません。微小な欠失を一部の施設がオプションで調べる場合もありますが、対象は限られ検出率にも幅があります。確定には羊水検査・絨毛検査と染色体マイクロアレイが必要です。
Q. 高い泣き声はずっと続きますか?
特徴的な高い泣き声は乳児期に目立ちやすく、成長とともに目立たなくなることが多いとされています。研究では、この泣き声が5p15.31付近の欠失と関連することが示されました。ただし現れ方には個人差があります。気になる泣き声が続くときは、自己判断せずかかりつけの小児科へ相談してください。
Q. 欠けた場所で症状は変わりますか?
患者94人を調べた研究では、高い泣き声は5p15.31付近、言葉の遅れは5p15.32〜15.33付近、顔立ちの特徴は5p15.2〜15.31付近の欠失と関連していました。知的発達の程度は、欠失の大きさと位置におおむね依存します。ただし、これは傾向であり、一人ひとりの見通しをそのまま決めるものではありません。
Q. どんな支援や療育が受けられますか?
乳幼児期からの理学療法・作業療法・言語の支援などが、発達をその子のペースで後押しします。合併症があれば各専門科が連携してケアします。難病情報センターや発達障害情報・支援センターなど、公的な相談窓口も心強い味方です。困ったときは一人で抱え込まず、専門職や地域の支援を頼ってください。
Q. 出生前に不安を感じたらどうすればよいですか?
まずは何をどこまで知りたいかを夫婦で話し合い、検査を段階的に考えることが助けになります。どの検査もすべての障害を見つけられるわけではなく、分かる範囲には限界があります。受ける・受けないは自己決定です。迷うときは遺伝カウンセリングで専門家に相談し、中立の情報をもとに落ち着いて判断してください。
著者・監修
岡 博史(おか ひろし)/医療法人社団福美会 ヒロクリニック統括院長・Labo Director(ラボディレクター)。慶應義塾大学医学部卒業後、日本と米国の医師国家試験に合格し、医学博士を取得。日本に約20人しかいないラボディレクター資格を保有。YouTube「ひろし先生の正しいエビデンス妊娠ch」などで、エビデンスにもとづく妊娠・出生前診断の情報を発信しています。
日本皮膚科学会 皮膚科専門医/日本医師会 産業医/東京衛生検査所 指導監督医
この記事は、 ヒロクリニックNIPTの編集・監修体制 にもとづき、資格を持つ医師が内容を確認しています。


